 W
WSexual selection is a mode of natural selection in which members of one biological sex choose mates of the other sex to mate with, and compete with members of the same sex for access to members of the opposite sex. These two forms of selection mean that some individuals have greater reproductive success than others within a population, for example because they are more attractive or prefer more attractive partners to produce offspring. For instance, in the breeding season, sexual selection in frogs occurs with the males first gathering at the water's edge and making their mating calls: croaking. The females then arrive and choose the males with the deepest croaks and best territories. In general, males benefit from frequent mating and monopolizing access to a group of fertile females. Females can have a limited number of offspring and maximize the return on the energy they invest in reproduction.
 W
WAnimal Coloration, or in full Animal Coloration: An Account of the Principal Facts and Theories Relating to the Colours and Markings of Animals, is a book by the English zoologist Frank Evers Beddard, published by Swan Sonnenschein in 1892. It formed part of the ongoing debate amongst zoologists about the relevance of Charles Darwin's theory of natural selection to the observed appearance, structure, and behaviour of animals, and vice versa.
 W
WThe Colours of Animals is a zoology book written in 1890 by Sir Edward Bagnall Poulton (1856–1943). It was the first substantial textbook to argue the case for Darwinian selection applying to all aspects of animal coloration. The book also pioneered the concept of frequency-dependent selection and introduced the term "aposematism".
 W
WThe Descent of Man, and Selection in Relation to Sex is a book by English naturalist Charles Darwin, first published in 1871, which applies evolutionary theory to human evolution, and details his theory of sexual selection, a form of biological adaptation distinct from, yet interconnected with, natural selection. The book discusses many related issues, including evolutionary psychology, evolutionary ethics, differences between human races, differences between sexes, the dominant role of women in mate choice, and the relevance of the evolutionary theory to society.
 W
WDisplay behaviour is a set of ritualized behaviours that enable an animal to communicate to other animals about specific stimuli. These ritualized behaviours can be visual however many animals depend on a mixture of visual, audio, tactical and/or chemical signals as well. Evolution has tailored these stereotyped behaviours to allow animals to communicate both conspecifically and interspecifically which allows for a broader connection in different niches in an ecosystem. It is connected to sexual selection and survival of the species in various ways. Typically, display behaviour is used for courtship between two animals and to signal to the female that a viable male is ready to mate. In other instances, species may exhibit territorial display behaviour, in order to preserve a foraging or hunting territory for its family or group. A third form is exhibited by tournament species in which males will fight in order to gain the 'right' to breed. Animals from a broad range of evolutionary hierarchies avail of display behaviours - from invertebrates such as the simple jumping spider to the more complex vertebrates like the harbour seal.
 W
WThe Evolution of Beauty: How Darwin's Forgotten Theory of Mate Choice Shapes the Animal World—and Us is a 2017 book by the ornithologist and evolutionary biologist Richard O. Prum about the power of aesthetic mate choice, arguing it to be an important independent agent in evolution. Prum indicates that while Charles Darwin made this argument in The Descent of Man, published in 1871, the concept was sidelined and forgotten and the notion of natural selection being the sole driver of evolution took over. As an ornithologist, Prum describes many examples in avian evolution where aesthetics are preeminent. Prum proceeds to apply the principle of aesthetic evolution as an independent force in human evolution.
 W
WThe evolution of sexual reproduction is an adaptive feature which is common to almost all multi-cellular organisms with many being incapable of reproducing asexually. Prior to the advent of sexual reproduction, the adaptation process whereby genes would change from one generation to the next happened very slowly and randomly. Sex evolved as an extremely efficient mechanism for producing variation, and this had the major advantage of enabling organisms to adapt to changing environments. Sex did, however, come with a cost. In reproducing asexually, no time nor energy needs to be expended in choosing a mate. And if the environment has not changed, then there may be little reason for variation, as the organism may already be well adapted. Sex, however, has evolved as the most prolific means of species branching into the tree of life. Diversification into the phylogenetic tree happens much more rapidly via sexual reproduction than it does by way of asexual reproduction.
 W
WThe theory of female cosmetic coalitions (FCC) represents a (controversial) attempt - proposed by evolutionary anthropologists Chris Knight and Camilla Power together with archaeologist Ian Watts - to explain the evolutionary emergence of art, ritual and symbolic culture in Homo sapiens.
 W
WFemale intrasexual competition is competition between women over a potential mate. Such competition might include self-promotion, derogation of other women, and direct and indirect aggression toward other women. Factors that influence female intrasexual competition include the genetic quality of available mates, hormone levels, and interpersonal dynamics.
 W
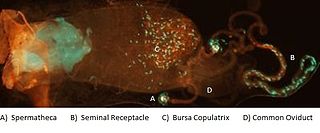
WFemale sperm storage is a biological process and often a type of sexual selection in which sperm cells transferred to a female during mating are temporarily retained within a specific part of the reproductive tract before the oocyte, or egg, is fertilized. The site of storage is variable among different animal taxa and ranges from structures that appear to function solely for sperm retention, such as insect spermatheca and bird sperm storage tubules, to more general regions of the reproductive tract enriched with receptors to which sperm associate before fertilization, such as the caudal portion of the cow oviduct containing sperm-associating annexins. Female sperm storage is an integral stage in the reproductive process for many animals with internal fertilization. It has several documented biological functions including:Supporting the sperm by: a.) enabling sperm to undergo biochemical transitions, called capacitation and motility hyperactivation, in which they become physiologically capable of fertilizing an oocyte and b.) maintaining sperm viability until an oocyte is ovulated. Decreasing the incidence of polyspermy. Enabling mating, ovulation and/or fertilization to occur at different times or in different environments. Supporting prolonged and sustained female fertility. Having a role influencing offspring sex ratios among some insects possessing a haplodiploid sex-determination system. Serving as an arena in which sperm from different mating males compete for access to oocytes, a process called sperm competition, and in which females may preferentially utilize sperm from some males over those of others, called female sperm preference or cryptic female choice.
 W
WFisherian runaway or runaway selection is a sexual selection mechanism proposed by the mathematical biologist Ronald Fisher in the early 20th century, to account for the evolution of exaggerated male ornamentation by persistent, directional female choice. An example is the colourful and elaborate peacock plumage compared to the relatively subdued peahen plumage; the costly ornaments, notably the bird's extremely long tail, appear to be incompatible with natural selection. Fisherian runaway can be postulated to include sexually dimorphic phenotypic traits such as behaviour expressed by either sex.
WFitness is the quantitative representation of natural and sexual selection within evolutionary biology. It can be defined either with respect to a genotype or to a phenotype in a given environment. In either case, it describes individual reproductive success and is equal to the average contribution to the gene pool of the next generation that is made by individuals of the specified genotype or phenotype. The fitness of a genotype is manifested through its phenotype, which is also affected by the developmental environment. The fitness of a given phenotype can also be different in different selective environments.
 W
WGenetic variance is a concept outlined by the English biologist and statistician Ronald Fisher in his fundamental theorem of natural selection which he outlined in his 1930 book The Genetical Theory of Natural Selection which postulates that the rate of change of biological fitness can be calculated by the genetic variance of the fitness itself. Fisher tried to give a statistical formula about how the change of fitness in a population can be attributed to changes in the allele frequency. Fisher made no restrictive assumptions in his formula concerning fitness parameters, mate choices or the number of alleles and loci involved.
 W
WGynomorph is a word used to describe an organism with female physical characteristics.
WThe handicap principle is a hypothesis proposed by Amotz Zahavi to explain how evolution may lead to "honest" or reliable signalling between animals which have an obvious motivation to bluff or deceive each other. It suggests that costly signals must be reliable signals, costing the signaller something that could not be afforded by an individual with less of a particular trait. For example, in sexual selection, the theory suggests that animals of greater biological fitness signal this status through handicapping behaviour, or morphology that effectively lowers this quality. The central idea is that sexually selected traits function like conspicuous consumption, signalling the ability to afford to squander a resource. Receivers then know that the signal indicates quality, because inferior quality signallers are unable to produce such wastefully extravagant signals.
 W
WIn population genetics, the Hardy–Weinberg principle, also known as the Hardy–Weinberg equilibrium, model, theorem, or law, states that allele and genotype frequencies in a population will remain constant from generation to generation in the absence of other evolutionary influences. These influences include genetic drift, mate choice, assortative mating, natural selection, sexual selection, mutation, gene flow, meiotic drive, genetic hitchhiking, population bottleneck, founder effect and inbreeding.
 W
WSperm competition is a form of post-copulatory sexual selection whereby male ejaculates simultaneously physically compete to fertilize a single ovum. Sperm competition occurs between sperm from two or more rival males when they make an attempt to fertilize a female within a sufficiently short period of time. This results primarily as a consequence of polyandrous mating systems, or due to extra-pair copulations of females, which increases the chance of cuckoldry, in which the male mate raises a child that is not genetically related to him. Sperm competition among males has resulted in numerous physiological and psychological adaptations, including the relative size of testes, the size of the sperm midpiece, prudent sperm allocation, and behaviors relating to sexual coercion, however this is not without consequences: the production of large amounts of sperm is costly and therefore, researchers have predicted that males will produce larger amounts of semen when there is a perceived or known increase in sperm competition risk. Sperm competition is not exclusive to humans, and has been studied extensively in other primates, as well as throughout much of the animal kingdom. The differing rates of sperm competition among other primates indicates that sperm competition is highest in primates with multi-male breeding systems, and lowest in primates with single-male breeding systems. Compared to other animals, and primates in particular, humans show low-to-intermediate levels of sperm competition, suggesting that humans have a history of little selection pressure for sperm competition.
 W
WHypergamy is a term used in social science for the act or practice of a person marrying a spouse of higher caste or social status than themselves.
 W
WKoinophilia is an evolutionary hypothesis proposing that during sexual selection, animals preferentially seek mates with a minimum of unusual or mutant features, including functionality, appearance and behavior. Koinophilia intends to explain the clustering of sexual organisms into species and other issues described by Darwin's Dilemma. The term derives from the Greek, koinos, "common", "that which is shared", and philia, "fondness".
 W
WA love dart is a sharp, calcareous or chitinous dart which some hermaphroditic land snails and slugs create. Love darts are both formed and stored internally in a dart sac. These darts are made in sexually mature animals only, and are used as part of the sequence of events during courtship, before actual mating takes place. Darts are quite large compared to the size of the animal: in the case of the semi-slug genus Parmarion, the length of a dart can be up to one fifth that of the semi-slug's foot.
 W
WThe major histocompatibility complex in sexual selection concerns how major histocompatibility complex (MHC) molecules allow for immune system surveillance of the population of protein molecules in a host's cells. In 1976, Yamazaki et al. demonstrated a sexual selection mate choice by male mice for females of a different MHC.
 W
WMate choice is one of the primary mechanisms under which evolution can occur. It is characterized by a “selective response by animals to particular stimuli” which can be observed as behavior. In other words, before an animal engages with a potential mate, they first evaluate various aspects of that mate which are indicative of quality—such as the resources or phenotypes they have—and evaluate whether or not those particular trait(s) are somehow beneficial to them. The evaluation will then incur a response of some sort.
 W
WMicrochimerism is the presence of a small number of cells that originate from another individual and are therefore genetically distinct from the cells of the host individual. This phenomenon may be related to certain types of autoimmune diseases; however, the mechanisms responsible for this relationship are unclear. The term comes from the prefix "micro" + "chimerism" based on the Chimera of Greek mythology.
 W
WNatural selection is the differential survival and reproduction of individuals due to differences in phenotype. It is a key mechanism of evolution, the change in the heritable traits characteristic of a population over generations. Charles Darwin popularised the term "natural selection", contrasting it with artificial selection, which in his view is intentional, whereas natural selection is not.
 W
WParental investment, in evolutionary biology and evolutionary psychology, is any parental expenditure that benefits offspring. Parental investment may be performed by both males and females, females alone or males alone. Care can be provided at any stage of the offspring's life, from pre-natal to post-natal.
 W
WReproductive success is defined as an individual's production of offspring per breeding event or lifetime. This is not limited by the number of offspring produced by one individual, but also the reproductive success of these offspring themselves. Reproductive success is different from fitness in that individual success is not necessarily a determinant for adaptive strength of a genotype since the effects of chance and the environment have no influence on those specific genes. Reproductive success turns into a part of fitness when the offspring are actually recruited into the breeding population. If offspring quantity is not correlated with quality this holds up, but if not then reproductive success must be adjusted by traits that predict juvenile survival in order to be measured effectively. Quality and quantity is about finding the right balance between reproduction and maintenance and the disposable soma theory of aging tells us that a longer lifespan will come at the cost of reproduction and thus longevity is not always correlated with high fecundity. Parental investment is a key factor in reproductive success since taking better care to offspring is what often will give them a fitness advantage later in life. This includes mate choice and sexual selection as an important factor in reproductive success, which is another reason why reproductive success is different from fitness as individual choices and outcomes are more important than genetic differences. As reproductive success is measured over generations, Longitudinal studies are the preferred study type as they follow a population or an individual over a longer period of time in order to monitor the progression of the individual(s). These long term studies are preferable since they negate the effects of the variation in a single year or breeding season.
 W
WA roar is a type of animal vocalization consisting of both a low fundamental frequency (pitch) and low formant frequency. Many mammals have evolved to produce roars and other roar-like vocalizations for purposes such as long-distance communication and intimidation. These include big cats, bears, some pinnipeds, hammer-headed bats, elephants, various bovids, red deer, gorillas and howler monkeys.
 W
WSecondary sex characteristics are features that appear during puberty in humans, and at sexual maturity in other animals. These characteristics are particularly evident in the sexually dimorphic phenotypic traits that distinguish the sexes of a species, but unlike the sex organs, are not directly part of the reproductive system. Secondary sex characteristics are believed to be the product of sexual selection for traits which display fitness, giving an organism an advantage over its rivals in courtship and in aggressive interactions.
 W
WSexual conflict or sexual antagonism occurs when the two sexes have conflicting optimal fitness strategies concerning reproduction, particularly over the mode and frequency of mating, potentially leading to an evolutionary arms race between males and females. In one example, males may benefit from multiple matings, while multiple matings may harm or endanger females, due to the anatomical differences of that species.
 W
WSexual dimorphism is the condition where the two sexes of the same species exhibit different characteristics beyond the differences in their sexual organs. The condition occurs in many animals and some plants. Differences may include secondary sex characteristics, size, weight, colour, markings, and may also include behavioral and cognitive differences. These differences may be subtle or exaggerated, and may be subjected to sexual selection and natural selection. The opposite of dimorphism is monomorphism.
 W
WSexual selection in amphibians involves sexual selection processes in amphibians, including frogs, salamanders and newts. Prolonged breeders, the majority of frog species, have breeding seasons at regular intervals where male-male competition occurs with males arriving at the waters edge first in large number and producing a wide range of vocalizations, with variations in depth of calls the speed of calls and other complex behaviours to attract mates. The fittest males will have the deepest croaks and the best territories, with females known to make their mate choices at least partly based on the males depth of croaking. This has led to sexual dimorphism, with females being larger than males in 90% of species, males in 10% and males fighting for groups of females.
 W
WSexual selection in Arabidopsis thaliana is a mode of natural selection by which the flowering plant Arabidopsis thaliana selects mates to maximize reproductive success.
 W
WSexual selection in birds concerns how birds have evolved a variety of mating behaviors, with the peacock tail being perhaps the most famous example of sexual selection and the Fisherian runaway. Commonly occurring sexual dimorphisms such as size and color differences are energetically costly attributes that signal competitive breeding situations. Many types of avian sexual selection have been identified; intersexual selection, also known as female choice; and intrasexual competition, where individuals of the more abundant sex compete with each other for the privilege to mate. Sexually selected traits often evolve to become more pronounced in competitive breeding situations until the trait begins to limit the individual's fitness. Conflicts between an individual fitness and signaling adaptations ensure that sexually selected ornaments such as plumage coloration and courtship behavior are “honest” traits. Signals must be costly to ensure that only good-quality individuals can present these exaggerated sexual ornaments and behaviors.
WSexual selection in humans concerns the concept of sexual selection, introduced by Charles Darwin as an element of his theory of natural selection, as it affects humans. Sexual selection is a biological way one sex chooses a mate for the best reproductive success. Most compete with others of the same sex for the best mate to contribute their genome for future generations. This has shaped our evolution for many years, but reasons why humans choose their mates are hardly understood. Sexual selection is quite different in non-human animals than humans as they feel more of the evolutionary pressures to reproduce and can easily reject a mate. The role of sexual selection in human evolution has not been firmly established although neoteny has been cited as being caused by human sexual selection. It has been suggested that sexual selection played a part in the evolution of the anatomically modern human brain, i.e. the structures responsible for social intelligence underwent positive selection as a sexual ornamentation to be used in courtship rather than for survival itself, and that it has developed in ways outlined by Ronald Fisher in the Fisherian runaway model. Fisher also stated that the development of sexual selection was "more favourable" in humans.
 W
WSexual selection in insects is about how sexual selection functions in insects. The males of some species have evolved exaggerated adornments and mechanisms for self-defense. These traits play a role in increasing male reproductive expectations by triggering male-male competition or influencing the female mate choice, and can be thought of as functioning on three different levels: individuals, colonies, and populations within an area.
 W
WSexual selection in mammals started with Charles Darwin's observations concerning sexual selection, including sexual selection in humans, and in other mammals, consisting of male-male competition and mate choice that mold the development of future phenotypes in a population for a given species.
 W
WSexual selection in scaled reptiles studies how sexual selection manifests in snakes and lizards, which constitute the order Squamata of reptiles. Each of the over three thousand snakes use different tactics in acquiring mates. Ritual combat between males for the females they want to mate with includes topping, a behavior exhibited by most viperids in which one male will twist around the vertically elevated fore body of its opponent and forcing it downward. It is common for neck biting to occur while the snakes are entwined.
 W
WThe sexy son hypothesis in evolutionary biology and sexual selection, proposed by Patrick J. Weatherhead and Raleigh J. Robertson of Queen's University in Kingston, Ontario in 1979, states that a female's ideal mate choice among potential mates is one whose genes will produce male offspring with the best chance of reproductive success. This implies that other benefits the father can offer the mother or offspring are less relevant than they may appear, including his capacity as a parental caregiver, territory and any nuptial gifts. Fisher's principle means that the sex ratio is always near 1:1 between males and females, yet what matters most are her "sexy sons'" future breeding successes, more likely if they have a promiscuous father, in creating large numbers of offspring carrying copies of her genes. This sexual selection hypothesis has been researched in species such as the European pied flycatcher.
 W
WWithin evolutionary biology, signalling theory is a body of theoretical work examining communication between individuals, both within species and across species. The central question is when organisms with conflicting interests, such as in sexual selection, should be expected to provide honest signals rather than cheating. Mathematical models describe how signalling can contribute to an evolutionarily stable strategy.
WSpeciation is the evolutionary process by which populations evolve to become distinct species. The biologist Orator F. Cook coined the term in 1906 for cladogenesis, the splitting of lineages, as opposed to anagenesis, phyletic evolution within lineages. Charles Darwin was the first to describe the role of natural selection in speciation in his 1859 book On the Origin of Species. He also identified sexual selection as a likely mechanism, but found it problematic.
WSperm competition is the competitive process between spermatozoa of two or more different males to fertilize the same egg during sexual reproduction. Competition can occur when females have multiple potential mating partners. Greater choice and variety of mates increases a female's chance to produce more viable offspring. However, multiple mates for a female means each individual male has decreased chances of producing offspring. Sperm competition is an evolutionary pressure on males, and has led to the development of adaptations to increase males' chance of reproductive success. Sperm competition results in a sexual conflict of interest between males and females. Males have evolved several defensive tactics including: mate-guarding, mating plugs, and releasing toxic seminal substances to reduce female re-mating tendencies to cope with sperm competition. Offensive tactics of sperm competition involve direct interference by one male on the reproductive success of another male, for instance by physically removing another male's sperm prior to mating with a female. For an example, see Gryllus bimaculatus.
 W
WSexual selection in spiders shows how sexual selection explains the evolution of phenotypic traits in spiders. Male spiders have many complex courtship rituals and have to avoid being eaten by the females, with the males of most species survive a few matings, and having short life spans.